| Great Lakes Ecosystems | ||
|
||||||||||
|
1994
Proceedings
|
|
SPATIAL DEMOGRAPHY OF AN OAK SAVANNA IN THE OZARKS
Sean E. Jenkins
|
 |
There is growing interest in how fire influenced the structure and composition of oak-dominated communities (savannas, glades and woodlands) and their distribution across the landscape mosaic. In eastern North America, oak ecosystems have evolved under the influence of repeated fire for thousands of years (Abrams 1992). However, fire suppression over the last 90 years has lead to an unprecedented conversion of savanna to forest with subsequent changes in overstory composition, structure, and loss of the diversity characteristic of savannas.
Several workers (Apfelbaum and Haney 1990; Guyette and Cutter 1991) have alluded to the existence of multiple even-aged cohorts in the overstory of Midwest savannas and hypothesized that fire-free periods are necessary for regeneration. However, little research has been done on spatial demographics of oaks and other species in relation to fire. Much of the present knowledge concerning the vegetation dynamics of savannas is based on small, remnant or partially restored sites, which may not be applicable to presettlement-, functional, savanna-oak woodland landscapes. To effectively manage oak ecosystems, information is needed concerning the relationships between fire history and oak regeneration at various spatial and temporal scales. In this study we examined how the historical fire regime has influenced the spatial demography of a continually-burned oak savanna in Camden County, Missouri. We were particularly interested in how oaks and other important species persist and are recruited into the overstory in response to very frequent fires.
The study site, AB Savanna, has an overstory typical of chert savannas in the Ozarks: post oak (Quercus stellata), white oak (Q. alba), black oak (Q. velutina), blackjack oak (Q. marilandica), and black hickory (Carya texana). However it is one of the few sites in the Midwest where fire has not been suppressed in this century.
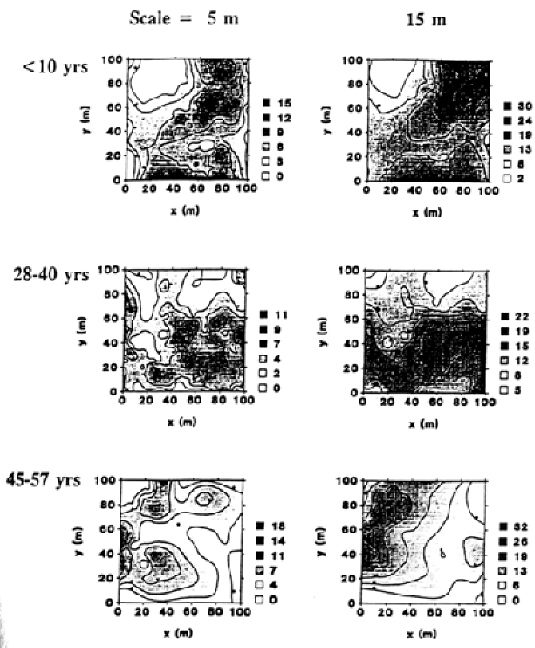
In 1993-1994, 977 stems >1.4 m tall within a 1-ha, west-facing plot were mapped and cored at 0.2-0.3 m above ground level. To supplement the core data, basal disks were cut from several hundred small-diameter stems that were killed in a fire, spring 1994. Complete ages were obtained from 95% of live stems. Ages of rotten trees were estimated from either 1) cores taken at 1.4 m height or 2) age-diameter regression equations. Spatial patterns of individual species and discrete cohorts (e.g., trees <10 years old) were analyzed using second-order neighborhood analysis (Getis and Franklin 1987) and plotted using the CONTOUR procedure in SYSTAT (Wilkinson 1989).
Tree ages ranged from 2 to 304 years. The age structures for the dominant tree species are all more or less continuous and multimodal, with at least four distinct cohorts (Fig. 1). With the exception of blackjack oak, all species were well represented by stems <10 years old. This class could be further resolved into 3-, 4-and 6-year-old cohorts, dating from spring fires in 1990, 1989, and 1987, respectively. However, at least some individuals of most species were recruited every year in the past decade. There has been very little new recruitment of blackjack oaks in the past 20 years. A small peak of trees 15 to 20 years old was recognizable for black hickory, black oak, and post oak. All species were represented by a cohort in the 30- to 40-year range. There were 120 black oaks/ha in this class, comprising a dominant patch feature in the savanna. Post oak and blackjack oak both had distinct cohorts in the 45- to 55-year-old range; this cohort was conspicuously absent in the other species. Post oak and white oak both had distinct cohorts in the 70- to 80-year range, a class absent or poorly represented in other species. The age class distributions- are more discontinuous for trees >100 years old. Older even-aged cohorts are not easily distinguishable, but a remnant multi-cohort pattern probably exists at a coarser scale.
Stem maps indicate that different species have congruent patterns for specific cohorts (i.e., species of similar age tend to be clumped together), although the youngest cohort (<10 years) of white oaks seems to be clustered around several large stump holes, which may have been old, veteran "wolf" trees. In contrast, different age cohorts tend to have complementary, non-overlapping patterns (Fig. 2). For example, most the new regeneration is concentrated in the northeastern quarter and extreme southern end of the plot. In contrast, the 28- to 40-year-old cohort is sandwiched in between the two younger cohort patches. Conversely, the 45- to 57-year-old cohort is most intensely clustered in the western part of the plot. The oldest trees (>135 years) were more randomly dispersed across the plot.
Although fire-free periods of several years may be responsible for some of the major even-aged cohorts present at AB Savanna, continuous regeneration also contributes substantially to the structure and dynamics. We are currently constructing a detailed fire history of the site to examine the influence of fire frequency (and intensity) on tree regeneration. We suspect that continuous regeneration is possible with annual or near-annual fires, which may reduce fuel loads and increase burn patchiness. Thus, there are both temporal and spatial "windows" for regeneration. However, these opportunities- to regenerate substantially vary among species. This is evident in the shifting dominance of certain cohorts among different species (Fig. 1). The younger cohorts are more dominant in black hickory, black oak, and blackjack oak; whereas, the older cohorts are more dominant in post oak. Neither post oak nor blackjack oak seem to regenerating well under the current, near-annual fire regime; however, low recruitment for post oak may be offset by its great longevity (i.e., k-selected species). Black hickory and black oak seem to persist with near-annual fires, but waves of regeneration accompany longer fire-free intervals when fuel loads buildup and the subsequent fire kills back the majority of stems.
Our results indicate that frequently-burned savannas have a complex spatial mosaic of even-aged patches, which probably develops over long periods under a variable fire regime. This variation not only affects patch distribution, but probably composition as well. Therefore, the systematic removal of smaller stems during restoration to capture a "snap-shot" of savanna structure in time may be altering the integrity of a system that developed gradually over millennia.
This research was funded by the USDA Forest Service, North Central Forest Experiment Station study "Assessing Impacts of Disturbance in Central Hardwood Ecosystems." We thank Richard Willis and his family for allowing us access to AB Savanna, for their friendship, and sharing their knowledge of fire lore in the Ozarks.
Apfelbaum, S. I. and A. W. Haney. 1990. Structures and Dynamics of Midwest Oak Savannas. IN: J. M. Sweeney, ed. Proceedings Symposium on Management of Dynamic Ecosystems. 51st Midwest Fish and Wildlife Conference. Springfield, IL.
Abrams, M. D. 1992. Fire and the development of oak forest. BioScience 42:346-353.
Getis, A. and J. Franklin. 1987. Second-order neighborhood analysis of mapped point patterns. Ecology 68: 473-477.
Guyette, R. P., and B. E. Cutter. 1991. Tree ring analysis of fire history of a post oak savanna in the Missouri Ozarks. Natural Areas Journal 11:93-99.
Wilkinson, L. 1989. SYGRAPH: The system for graphics. Systat, Inc., Evanston, IL.
Figure 1. Age frequency distributions of the major overstory species at AB Savanna
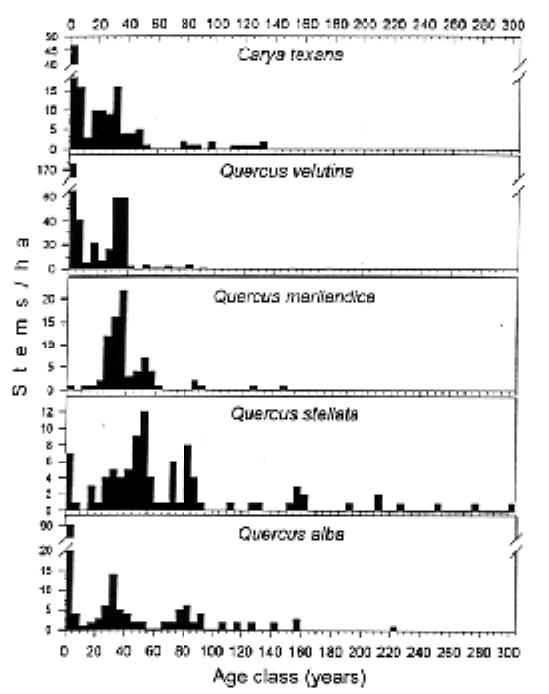
Figure 2. Spatial patterns of three cohorts at scales of 5 and 15 m. L(d) values for individual trees are interpolated to create a contour map of clumping intensity. L(d) values increase with increasing clumping intensity and are proportional to the number of neighborhood trees around each tree at the specified scale or radius.