| Great Lakes Ecosystems | ||
|
||||||||||
|
1994
Proceedings
| ||||||||||||||||
|
POLLEN ANALOG DATES FOR MIDWESTERN OAK SAVANNAS
Duane Griffin
|
 |
Most paleoecological reconstructions of North American vegetation fail to recognize transitional vegetation types such as oak savannas. Consequently, we know little about their ecological histories. One approach to reconstructing these histories is by extrapolation from mapped summaries. Isopoll (contours of constant pollen percentage) maps in Webb (1988) show southern Wisconsin and adjacent areas as characterized by high percentages of oak and grass pollen and high to moderate prairie forb percentages at 500 years BP. By overlaying maps for these taxa, it appears that similar combinations of taxa may have occurred as early as 10000 BP in eastern Nebraska and migrated eastward. Webb's maps, however, are very small scale, with coarse spatial and temporal resolution.
Another approach is the analog method of pollen analysis (Overpeck et al. 1985), wherein modern pollen spectra are compared to the fossil record to identify modern analogs to past vegetation. In this study I identify and date the origin of the oak openings (Curtis 1959) by comparing presettlement (PS) pollen spectra from southern Wisconsin and northern Illinois with the fossil pollen record from twenty six sites in the upper Midwest.
Oak savannas of various types occurred from Texas to Minnesota until the 19th century (Nuzzo 1985). Curtis (1959) delimited oak openings as communities dominated by grasses having > 1 tree/acre and less than 50% canopy cover, most commonly bur oak (Quercus macrocarpa). Oak openings were most extensive in southern Wisconsin and extended into northeastern Illinois and southeastern and central Minnesota (hereafter referred to as the Wisconsin savanna region). In Iowa and central Illinois the prairie-forest ecotone seems to have been more of a mosaic of lowland forests and upland prairies than a composite of forest and prairie elements as interpreted by Curtis (1959; c.f. Davis 1977; Nuzzo 1985).
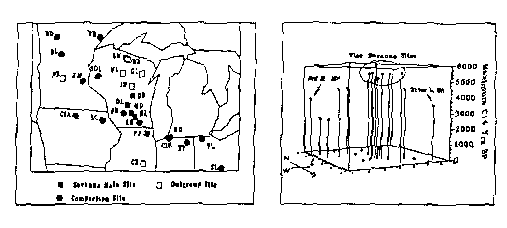
Using data files provided by L.J. Maher, I selected 26 sites for analysis (Fig. 1) and isolated 19 pollen taxa, combined into ten groups1. Taxa selection followed Curtis (1959) with the addition of herbaceous pollen. I interpolated ages between radiocarbon dates for each level of 26 of the cores using SLOTDEPV (Maher 1991). Five sites indicated as oak openings on the WGNHS (1965) map form the main savanna group. Lima Bog, WI is in the PS savanna region but is not included because of anomalously high grass pollen sums, probably from wild rice (Baker et al 1992). Nine sites from conifer and mesic deciduous forests and prairie regions are included as an outgroup. I isolated the pollen spectra from levels just below the rise in Ambrosia pollen, which marks postsettlement land clearing, from these files and combined them into a single data file, structured as if it were a pollen core.
I used SLOTDEPV (Maher 1991) to produce correlation matrices for various chord distance dissimilarity coefficients (Overpeck et al. 1985). Dissimilarity coefficients measure differences between multivariate samples; similar pollen spectra produce low dissimilarity values, suggesting that similar vegetation produced them. SLOTDEPV calculates the dissimilarity between each level of two cores and displays the results graphically. By varying the maximum value displayed, I determined that CD = 0.264 effectively discriminated the oak opening spectra from other vegetation types. Finally, I compared the dummy core to each of the 26 cores and recorded the earliest match between PS savanna and fossil spectra. I considered a match to be valid when three or more contiguous levels in the core matched at least one PS spectrum. _______________________________________________
Oak opening spectra are characterized by higher oak percentages than prairie sites, and higher herbaceous and lower hardwood percentages than forest sites. Figure 2 shows the maximum ages for analogs to the PS oak opening pollen spectra. The oldest dates, 5500-6000 radiocarbon years BP, come from the Wisconsin sites. The 6000 BP date is from Lake Mendota, WI, a carbonate lake, and is probably too old. The other Wisconsin savanna dates (5500 BP) match those obtained at Devil's Lake, WI, which is situated in crystalline bedrock and thus has very good dating control. Clear Lake, IN, matched the PS spectra for four of the main savanna group sites from 10000-8000 BP. I interpret this as a spurious match, since the spectra at Hudson Lake, MI and other sites in the region reflect oak-hardwood forests.
The Iowa sites generated no matches with the Wisconsin savanna sites, contradicting the working hypothesis of western origin and eastward migration. The oak openings seem to have originated in situ in S. Wisconsin and N Illinois 5500 BP, a time when the climate of the region became warmer and drier (Baker et al. 1992; Winkler et al. 1986). At that time, the ecotone between Wisconsin's northern and southern vegetation types shifted northwards and disturbance regimes on both sides of it shifted from low to high disturbance (Griffin 1994). In southern Wisconsin, the mesic forests that had dominated the region since ca. 9500 BP suddenly disappeared, giving rise to the oak openings. This warm and dry period began much earlier in Minnesota, where McAndrews (1966) identified oak savanna as early as 8500 BP. The lack of a match between the PS savanna spectra and the fossil record from McAndrews' Bog D until 4000 BP suggests that the early and later savannas were different.
The 5500 BP date for the savanna origin is conservative. It is limited by the low level of taxonomic resolution available from pollen data, rendered even lower by consideration of only 10 taxa groups, but it is appropriate to the level of generalization of the WGNHS map to which it is tied. Finer discrimination in delimiting vegetation types would doubtlessly reveal the savannas that existed from 5500-3500 BP to be different from those that existed from 3500 BP (when cooler and moister conditions resumed) until the 19th century. At even finer resolution, species composition and abundance in the oak openings might well reflect the effects of the Little Ice Age (ca 1500-1900 AD).
If correct, these findings have two important implications for oak savanna restoration. First, they suggest that evolutionary rationales for restoration (and strategies based on them) may be misguided. In a debate on the validity of savanna restoration, Packard (1993) argues that savannas are legitimate foci for restoration efforts because even transitional vegetation types represent "complex assemblages of species as they have evolved in their environments over the ages." Indeed, this is an underlying rationale for virtually all scientific restoration (sensu Howell 1988). On the other hand, Davis (1981) cautions that, before making inferences about interspecific adaptations, we should "inspect the paleoecological record...to be sure that...species have co-occurred long enough for evolutionary adjustments to have taken place." Unless we are very wrong in our understanding of evolutionary rates, 5000 years is scarcely enough time for evolution to have occurred.
Second, the savanna species (apparently) organized themselves into viable communities rather rapidly in response to unique environmental conditions. Restoring those conditions should, likewise, create viable, though certainly different, savannas. Realistically, this is the only option available, since the conditions (including climate) that fostered the original savannas have changed forever, and we lack sufficient information on their composition to attempt accurate recreations. This approach to restoration promises its own scientific rewards which, along with aesthetic and historical considerations, should provide adequate justification for the effort. We need not, and should not, appeal to spurious arguments invoking evolution where none has occurred.
Lou Maher (UW Madison Department of Geology and Geophysics) provided the raw data files and analytical software used in this study.
Baker, R.G., L.J. Maher, C.J. Chumbley, and K.L. Van Zant. 1992. Patterns of Holocene environmental change in the Midwestern United States. Quaternary Research 37:379-389.
Curtis, J.T. 1959/1987. The Vegetation of Wisconsin. Madison: Univ. of Wisconsin Press.
Davis, A.M. 1977. The prairie-deciduous forest ecotone in the upper middle west. Annals of the Association of American Geographers 67:204-213.
Davis, M.B. 1981. Quaternary history and the stability of forest communities. In D.C. West, H.H. Shugart, and D.B. Botkin, eds. Forest Succession: Concepts and Applications. New York: Springer Verlag.
Griffin, D.A. 1994. Disturbance and Ecotone Dynamics in the Holocene Vegetation History of Wisconsin. Paper presented at the 90th Annual Meeting of the Association of American Geographers, San Francisco, California.
Howell, E. 1988. The role of Restoration in Conservation Biology. Endangered Species Update 5(3&4):1-5)
McAndrews, j.H. 1966. Postglacial history of prairie, savanna and forest in northwestern Minnesota. Memoirs of the Torrey Botanical Club 22: 1-72.
Maher, L.J. 1991. DEP-AGE (Version 3.2), PLOTSITE (Version 2.8), POLFILE (Version 1.15), and SLOTDEPV (version 1.2) computer programs for pollen analysis. Univ. of Wisconsin Department of Geology and Geophysics.
Nuzzo, V.A. 1985. The extent and status of Midwest oak savanna at the time of settlement and in the mid 1980's... M.S. Thesis, Univ. of Wisconsin, Madison.
Overpeck, J.T., T. Webb III, and I.C. Prentice. 1985. Quantitative interpretation of fossil pollen spectra: Dissimilarity coefficients and the method of modern analogs. Quaternary Research 23:87-108.
Packard, S. 1993. Oak savanna restoration. Restoration and Management Notes 11:5-1.
Webb, T. III. 1988. Eastern North America. In B. Huntley and T.Webb III (eds), Vegetation History, pp. 385-414, Dordrecht: Kluwer Academic Publishers.
Webb, T. III, E.J. Cushing, and H.E. Wright, Jr. 1983. Holocene changes in the vegetation of the Midwest. In: H.E. Wright, Jr, ed. Late Quaternary Environments of the United States, Vol. 2, The Holocene. Minneapolis: Univ. of Minnesota Press.
Winkler, M.G., A.M. Swan, and J.E. Kutzbach. 1986. Middle Holocene dry period in the Northern Midwestern United States: Lake Levels and Pollen Stratigraphy. Quaternary Research 25:235-250.
WGNHS. 1965. Early Vegetation of Wisconsin (Map). Madison: Wisconsin Geological and Natural History Survey.
 |
|
|
Figure
1. General location of sites. 2 savanna |
Figure 2. Maximum ages of presettlement oak analogs. |
| 2-Site Key for Figure 1.
BL Billy's Lake (1); BM
Blue Mounds Cr. (1); BD Bog D (1); CB Chatsworth Bog (1); CIA Clear
Lake (2); CIN Clear Lake (1); DL Devils Lake (2); DB Disterhaft Farm
Bog (2); FL French Lake (1); GL Green Lake (A.M. Swain unpub.);
HK
Hells Kitchen Lake (1); HL Hook Lake (Winkler 1985 and Maher unpub.);
HD
Hudson Lake (1); IB Iola Bog (2); KM Kirchner Marsh (1); LM Lake Mary
(1); MD Lake Mendota (1); LB Lima Bog (2; grass not included);
PL
Pretty Lake (1); RC Roberts Creek Sites (2); SL Silver Lake (1);
SDL
Stewart's Dark Lake (1); ST Stotzel-Leis Site (1); VB Volo Bog (1);
WB Weber
Lake (1); WL Wood Lake (1).
Citation Key: 1 = Webb et al 1983; 2 = Baker et al. 1992. |
|