| Great Lakes Ecosystems | ||
|
||||||||||
|
1994
Proceedings
|
|
ANALYSIS OF A FLOWER COLOR POLYMORPHISM IN VIOLA
PEDATA (BIRDFOOT VIOLET)
Steven B. Carroll |
 |
Viola pedata (birdfoot violet) is a common spring wildflower of prairie/forest edges and open woodlands throughout the eastern U.S. (Fernald 1970). In contrast to most violets, and indeed, to most flowering plants, V. pedata includes two plant types that substantially differ in flower color. One type (var. bicolor; hereafter Dark, or DK) produces striking bicolored flowers in which the two upper petals are dark violet and the three lower petals are lilac in color. In the other type (var. lineariloba; hereafter Light, or LT), all five petals are lilac. In our area, proportions of the two plant types vary from population to population but are relatively constant from year to year within populations.
We studied the reproductive ecology of this species in an attempt to answer the following questions: 1) What variation exists among populations with respect to proportions of the two morphs?, 2) Do pollinators discriminate against one or the other of the two morphs?, and 3) Do the two morphs differ with respect to fitness-related traits such as the numbers of flowers, ovules, fruits, and seeds produced?
Studies were done in several populations located in northeastern Missouri (Table 1), with sites concentrated in two clusters. Six northern sites were within 5 km of each other, just southwest of the city of Kirksville, MO. Four southern sites were located 30 km to the south at the perimeter of the Atlanta Wildlife Area; these sites were within 1 km of each other. One additional site (Mbly) was located 50 km south of these four sites.
Morph proportions were determined either by censusing the nearest individual at fixed intervals along transects (Table 1: 220C, Park, Atl1, Atl 2, Atl3a, Atl3b, Mbly) or by censusing all plants within a prescribed area (BC1, BC2, SC1, SC2).
A total of 649 insect visitors was observed. For each of these, a "flight path" was documented that included each visited flower and the plant type (DK or LT) for each. For 54 of these visitors, time spent on individual flowers was also recorded.
Reproductive allocation was quantified as the number of flowers/plant, ovules/ovary, fruits/plant, fruits/flower (fruit set), and seeds/fruit. Flowers and fruits were counted in the field; ovules and seeds were counted in the lab. Flowers were collected after anthesis. Fruits, which disperse seeds ballistically, were collected before dehiscence and allowed to open in coin envelopes.
Populations fell into two groups -- six northern populations in which DKs were more common, and five southern populations in which LTs were more common (Table 1). Morph proportions did not change significantly from year to year within sites. In addition, no plant was observed to change flower color between years.
Of 54 pollinators at four sites, 37 (69 %) were butterflies (36 of which were dusky wings, genus Erynnis), eight (15 %) were bees, four (7 %) were flies, three (5 %) were moths, one (2 %) was a wasp, and one (2 %) was unidentified. Dusky wings visited a total of 138 flowers (102 DK, 36 LT) on 107 plants (77 DK, 30 LT). At three of four sites, insects visited the two morphs according to their proportions in the population, indicating a lack of discrimination against either morph. Only at site Atl3a were the proportions of visited plants marginally significantly different from site proportions (X2 = 3.648, df = 2, P = 0.056); in this population, DKs were visited at a slightly higher proportion than expected. For an additional 595 flower visits at two DK-dominated sites, the proportions of visited flowers and the proportions of the two morphs in the populations did not differ statistically.
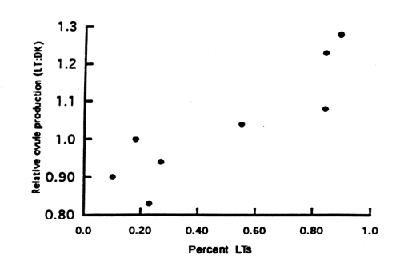
Mean numbers of flowers/plant, ovules/ovary, fruits/plant, fruits/flower, and seeds/fruit all varied significantly among sites in at least one year of the study. LT plants produced more flowers than DK plants in six of eight cases, but between-morph differences were statistically significant only at the Park site. The two sites at which DKs produced more flowers than LTs (BC1 94 and Park 94) were both DK-dominated sites. DKs produced more ovules/ovary at three of five DK-dominated sites, whereas LTs produced more ovules/ovary at all LT-dominated sites. At LT-dominated sites, both morphs - but especially DKs - produced fewer ovules/ovary. The Spearman rank correlation coefficient between relative ovule production [(number of ovules/ovary produced by LTs) / (number of ovules/ovary produced by DKs)] and the proportion of LTs in a population was highly statistically significant (r = 0.875, n = 8, P = 0.007) (Fig. 1).
DKs produced more fruits/plant in six of nine cases, three at DK-dominated sites and three at LT-dominated sites. However, none of the between-morph differences were statistically significant (n = 9). Between-morph fruit set differences were also statistically non-significant in all cases (n = 7). LTs produced greater numbers of seeds/fruit at seven of nine sites, but between-morph differences within sites were statistically significant only at BC1 in 1993. A measure of maximum potential seed production for individual plants can be expressed as: (# flowers/plant)(# ovules/flower). Relative potential seed production of LT with respect to DK averaged 1.03 at five DK-dominated sites, but averaged 1.63 at two LT-dominated sites. The Spearman rank correlation coefficient for the relationship between relative potential seed production and the percent of LTs in populations was marginally significant (r = 0.750, n = 7, P = 0.053).
The proportions of DKs and LTs varied widely among our 11 sites in northeast Missouri. Regional floras (e.g., Fernald 1970) indicate that the range of LT is greater than that of DK, suggesting that selection acts differently on the two morphs, at least in some parts of the range. This difference raises the interesting question of how the two morphs are able to co-occur in our populations, and throughout much of the range of the species. Assuming that flower color is under genetic control, even small differences in fitness-related traits should eventually lead to populations in which the proportion of the less successful morph is reduced to low mutation-selection levels (Futuyma 1986).
Pollinators have been shown to favor one color over another in other species with flower color polymorphisms (Kay 1976; Stanton and Preston 1987), and fitness is known to vary among flower-color morphs (Waser and Price 1981; Stanton et al. 1986). Fitness differences also have been found where pollinators do not discriminate between color morphs (Wolfe 1993). In any event, pollinators are generally expected to favor more common morphs, and in such a case, the less common morph would be expected to be eliminated. In our case, results are not clearcut. Between-morph differences were statistically significant at only one site for flower and seed production, at two sites for ovule production, and at no sites for fruit production. These results point out the value of conducting ecological studies in as many sites and years as possible. For example, at BC1, LTs produced significantly more seeds/fruit than DKs in 1993 but not in 1994. Spring 1993 was relatively cool and wet compared to spring 1994. Such differences can be expected to affect plant growth and pollinator activity, either or both of which may have contributed to between-year differences in our study.
One especially intriguing result was a statistically significant correlation between relative ovule production and the proportion of LTs in populations. This suggests at least the possibility that LTs eventually become dominant on sites at which they have the potential for higher fecundity, even if this potential is not realized in a particular year. There also exists the possibility of balancing selection acting on seed production. For example, at Atl 3a, where relative ovule/ovary of LT:DK was 1.04, pollinators favored DKs slightly over LTs. Although butterflies were the major insect visitors at our sites, Beattie (1976) reported solitary bees as the major visitor in West Virginia. Additional observations of pollinators under more varied conditions may help clarify their relative importance as selective forces on fitness in this species.
We thank Tracey Lassere and Becky Franklin for assistance with all aspects of this study, and NMSU for a summer faculty research grant.
Beattie, A. J. 1976. Plant dispersion, pollination and gene flow in Viola. Oecologia 25:291-300.
Fernald, M. L. 1970. Gray's Manual of Botany. Van Nostrand Reinhold Co., NY.
Futuyma, D. J. 1986. Evolutionary Biology. Sinauer Associates, Sunderland, MA.
Kay, Q. O. N. 1976. Preferential pollination of yellow-flowered morphs of Raphanus raphanistrum by Pieris and Eristalis spp. Nature 261:230-232.
Stanton, M. L. and R. E. Preston. 1987. The reproductive biology of petal color variants in wild populations of Raphanus sativus L.: I. Pollinator response to color morphs. American Journal of Botany 74:178-187.
Stanton, M. L., Snow, A. A., and S. N. Handel. 1986. Floral evolution: attractiveness to pollinators influences male fitness. Science 232:1625-1627.
Waser, N.M. and M.V. Price. 1981. Pollinator choice and stabilizing selection for flower color in Delphinium nelsonii. Evolution 35:376-390.
Wolfe, L. M. 1993. Reproductive consequences of a flower color polymorphism in Hydrophyllum appendiculatum. American Midland Naturalist 129:405-408.
Table 1. Site names, descriptions, density of all vegetation, and morph proportions for six northern, DK-dominated populations and five southern, LT-dominated populations. Northern and southern populations are approximately 30 km apart.
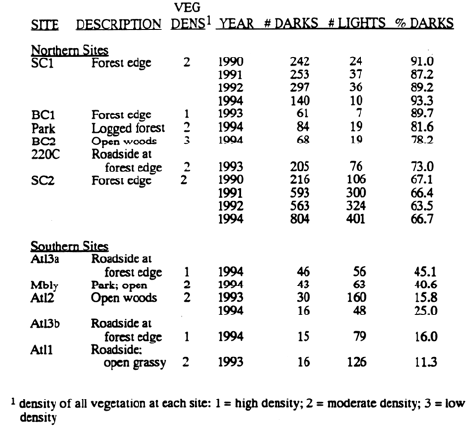
Figure 1. Relationship between relative ovule production (LT:DK) and the proportion of LTs in eight populations.